Il Fusto
Il fusto funzioni e struttura
Nelle cormofite, la presenza di tessuti
conduttori e di sostegno, ha reso
possibile lo sviluppo di un organo
epigeo, il fusto o caule che, assolve a
numerose funzioni:
1. Sostiene le porzioni fotosintetizzanti,
rappresentate in gran parte dalle foglie
e le espone alla luce nella giusta
angolazione;
2. Garantisce il trasporto delle sostanze
nutritive dalle radici e dalle foglie a tutte
le cellule della pianta;
3. Costituisce un organo di riserva, seppure
in misura molto minore rispetto alle
radici.
Il fusto presenta due tipi di
accrescimento:
• L’accrescimento primario
si origina dai
meristemi apicali dei germogli e delle radici.
L’accrescimento primario porta ad un
allungamento in lunghezza.
• L’accrescimento secondario
deriva
dall’accrescimento secondario o laterale, il
cambio vascolare o il cambio del sughero.
L’accrescimento secondario porta ad un
aumento di spessore.
Osservando l’estremità del fusto è possibile mettere
in evidenza tre zone che dall’apice, verso il basso
sono dette rispettivamente:
1 Zona di accrescimento per divisione
Le cellule del meristema apicale sono in continua divisione,
vi è produzione di nuovi elementi che allontanandosi dal
meristema, cessano di dividersi e vanno incontro ad un
processo di distensione e differenziazione.
2 Zona di accrescimento per distensione e di
differenziazione
È la zona in cui avviene l’allungamento del giovane fusto
3 Zona di struttura primaria
Dove le cellule sono ormai morfologicamente e
funzionalmente adulte.
L’accrescimento primario in lunghezza è a questo punto
terminato.
Struttura primaria del fusto
Effettuando una sezione trasversale di un
giovane fusto di dicotiledone ad una
certa distanza dall’apice (a livello della
struttura primaria), è possibile osservare
al microscopio ottico, dall’esterno verso
l’interno
• Epidermide
• Corteccia
• Cilindro centrale.
L’epidermide è lo strato più esterno
del fusto.
La corteccia è formata da più di un
tipo di tessuto. Negli strati più
superficiali, nei quali la radiazione
solare
penetra
facilmente,
è
presente un parenchima clorofilliano;
più all’interno, dove la radiazione
solare non arriva, i plastidi si saranno
differenziati in leucoplasti e il
parenchima che li contiene sarà
essenzialmente un parenchima di
riserva.
Nella corteccia sono presenti anche
tessuti meccanici (collenchimi e/o
sclerenchimi).
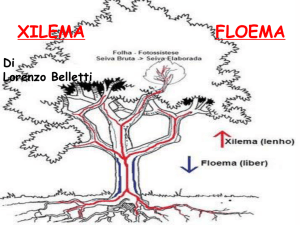
Il cilindro centrale è la zona più
interna del fusto in cui si trovano i
tessuti conduttori: i fasci (Xilema e
Fluoema).
La struttura primaria del fusto
Nelle spermatofite possiamo individuare tre tipi di struttura primaria.
Sifonostele
Il sistema vascolare
appare come un
cilindro intorno al
midollo.
Questa
struttura è tipica di
alcune conifere e
magnolidi.
Eustele
I fasci conduttori
appaiono distinti e
disposti in maniera
circolare intorno al
midollo.
Atactostele
I fasci conduttori
appaiono sparsi in
tutto
il
tessuto.
Questa struttura è
tipica
delle
monocotiledoni e delle
eucotiledoni
(classe
delle angiosperme)
Sifonostele
La sezione trasversale del fusto di Tilia
mostra la struttura sifonostelica dove la
corteccia ed il midollo sono separati
dall’anello dei tessuti conduttori.
La sezione trasversale del fusto di Medicago
mostra la struttura eustelica dove i fasci sono
separati fra loro. Il fusto di questa pianta
presenta anche una crescita secondaria.
Il
cambio
cribro-legnoso
è
in
parte
intrafascicolare, procambiale, ed in parte è
interfascicolare, parenchima interfascicolare.
Durante l’accrescimento secondario i
tessuti secondari si formano per la
maggior parte dalle cellule cambiali. Il
cambio interfascicolare produce solo
cellule sclerenchimatiche ai lati dello
xilema.
Eustele
La sezione trasversale all’altezza dell’internodo
del fusto di Zea mays mostra i fasci conduttori
disposti
irregolarmente
nel
tessuto
fondamentale. Questa
Struttura è tipica delle
monocotiledoni.
Atactostele
Connessione del sistema conduttore
con quello della foglia
I cordoni del procambio del fusto si
originano sotto i primordi fogliari. Pian
piano che la foglia si sviluppa i cordoni
procambiali del fusto si allungano nei
primordi fogliari. Si ha che il procambio
della foglia è continuo con quello del
fusto.
I fasci presenti nel cilindro centrale
attraversano la corteccia e entrano
nella foglia, all’atezza del nodo,
irrorandola. I prolungamenti dei fasci
prendono il nome di tracce fogliari.
Le zone sovrastanti le tracce fogliari
prendono il nome di lacuna fogliare.
Il fascio conduttore insieme alle sue
tracce fogliari prendi il nome di
simpodio.
L’accrescimento secondario
Perché è necessario che la pianta abbia questo tipo di accrescimento
secondario?
Necessità primaria di una pianta è quella di svolgere un’efficiente
attività fotosintetica, che richiede l’allungamento del fusto in
maniera da portare le foglie sempre + in alto.
Vi sarebbero problemi di staticità se non intervenisse l’accrescimento
in spessore, che permette anche lo sviluppo di nuovi fasci
conduttori in collegamento con le foglie neoformate.
Nelle gimnosperme e nelle dicotiledoni l’accrescimento secondario in
spessore è assicurato dall’attività di due meristemi:
1. Cambio cribro vascolare, che porta all’accrescimento del tessuto
vascolare
2. Cambio subero fellodermico, che origina un nuovo tessuto di
rivestimento, il periderma.
L’accrescimento secondario
Una volta che il fusto primario
ha raggiunto la maturazione, i
suoi tessuti sono costituiti da
cellule differenziate e adulte, le
quali non sono più in grado di
rigenerarsi per far fronte ad
un’ulteriore
crescita
degli apici.
A questo punto l’unico
modo che ha la pianta
per mantenere in vita gli apici e
farne crescere di nuovi, è quello
di costruire una nuova struttura:
alcune cellule adulte del fusto
regrediscono
allo
stato
embrionale e vanno a formare un
meristema
secondario,
così
chiamato perché non derivato
direttamente da quello primario,
ma appunto da cellule adulte.
Negli spazi compresi tra i fasci conduttori primari, si forma così uno strato di
cellule detto “cambio”, che si chiude attorno al fusto formando una
circonferenza. Questa in realtà si estende per tutta l’altezza del fusto, e forma
quindi un cilindro cavo di cellule meristematiche.
Il cambio duplica le proprie cellule, sia verso l’esterno, formando un anello
continuo di floema (libro), sia verso l’interno formando un anello di xilema
(legno): questi nuovi tessuti garantiscono la continuità degli scambi tra le
radici
e
gli
apici
in
continuo
accrescimento.
Via via che nuovi germogli si formano, i rami precedenti lignificano e
iniziano l’accrescimento secondario. Quindi con il passare del tempo, nei
rami più vecchi e nel tronco, il cambio deve continuare a produrre
internamete nuovo xilema che si sovrappone a quello più vecchio via via che
questo si degrada e perde la sua funzione. Le cellule dello xilema quando
muoiono rimangono nella parte più interna del fusto, assumendo funzione di
sostegno.
Esternamente al cambio avviene invece la formazione di nuovo floema.
In questo caso, l’espansione del cambio fa sì che il floema più recente
spinga il più vecchio verso l’esterno. La sua capacità di espandersi è
però limitata. Il floema più esterno si de-differenzia nuovamente,
creando un ulteriore tessuto meristematico, il fellogeno: questo va a
formare delle cellule tegumentarie: il periderma e la corteccia in cui
la cui parete cellulare è composta da sughero.
Il fellogeno ha vita breve: trovandosi in una zona non più alimentata,
le cellule della corteccia muoiono, e vengono spinte verso l’esterno,
deformandosi a causa della crescita, assumendo così un aspetto
fessurato.
Ogni anno quindi un nuovo strato di fellogeno si forma più internamente a partire
dal floema.
La maggior parte delle piante che vivono in climi temperati freddi, ha ritmi di
crescita che seguono le stagioni e sono perciò discontinui: durante la stagione
invernale la crescita subisce un arresto, per poi riprendere in primavera.
La crescita primaverile è quella che richiede più acqua, grazie alla formazione di
nuovi germogli e nuove foglie, per tanto lo xilema formato in primavera è
costituito da cellule molto più grandi e con parete cellulare sottile, che
consentono un maggior trasporto idrico, mentre durante l’estate la nascita di
nuove foglie rallenta, e di conseguenza le cellule xilematiche estive sono più
sottili, e con parete cellulare più spessa, con funzione di sostegno.
Terminata la stagione vegetativa la pianta va a riposo e smette di produrre nuovo
legno fino alla primavera successiva: questa alternanza annuale tra xilema
primaverile ed estivo è ben visibile a occhio nudo in una sezione trasversale del
tronco e consente di valutare l’età di un albero dal conteggio del numero di
cerchie annuali.
La struttura secondaria è tipica delle gimnosperme (conifere) e delle
angiosperme dicotiledoni (altre specie a fusti legnosi).
La classe delle angiosperme monocotiledoni invece è caratterizzata
dall’avere solo una struttura primaria: essa comprende per lo più piante
erbacee a ciclo annuale o perenne, bulbose, rizomatose, e anche piante simili
ad alberi, come le palme, caratterizzate dall’avere un unico apice primario
che si allunga senza avere accrescimento diametrale.
Cambio cribro vascolare
Sia nel fusto che nella radice, il cambio cribro vascolare è costituito da
un anello continuo di due tipi di cellule :
1. Iniziali fusiformi che sono allungate longitudinalmente,
appiattite ed affusolate alle estremità e per mezzo di divisioni
periclinali aumentano gli strati di cellule che formano il corpo
della pianta.
Quando una si divide, una delle due cellule figlie, dopo aver prodotto per
mitosi un numero variabile di derivate, si differenzia, mentre l’altra
cellula figlia rimane meristematica ed in grado di dividersi ancora.
2. Iniziali dei raggi, sono molto più corte delle iniziali fusiformi, si
dividono e producono raggi midollari.
Questi sono costituiti da cellule vive parenchimatiche che hanno
funzione di riserva e di trasporto delle sostanze nutritive.
Il numero delle derivate xilematiche è in genere maggiore di quello
delle derivate floematiche e ciò fa si che dal cambio si origini
molto + xilema secondario verso l’interno ed un quantitativo
inferiore di floema secondario verso l’esterno.
Le
cellule disposte internamente al cambio cribrovascolare si differenziano in tracheidi, elementi delle
trachee o altre cellule dello XILEMA (o legno)
SECONDARIO, mentre quelle esterne si differenziano
in cellule cribrose o elementi dei tubi cribrosi, cellule
annesse o altre cellule del FLOEMA (o libro)
SECONDARIO.
Cosi lo xilema secondario si trova in contatto con quello
primario, più interno, e il floema secondario è in
contatto con il primario più esterno. Questa è detta
attività dipleuretica
Cambio subero-fellodermico
A
differenza del cambio cribrovascolare il fellogeno è sempre in
tutta la sua estensione, un
meristema di origine secondaria in
quanto deriva da cellule già
differenziate che riacquistano le
caratteristiche meristematiche.
L’attività del fellogeno permette di
sostituire i tessuti primari di
rivestimento di fusto e radice, che
a causa della dilatazione tendono a
lacerarsi,
con
un
tessuto
secondario: il PERIDERMA.
Il fellogeno (≠ dal cambio cribro
vascolare) è formato da un solo
tipo di cellule iniziali, ma anch’esso
presenta attività dipleurica, in
quanto dalla divisione delle sue
cellule origina sughero(+) verso
l’esterno e felloderma(-) verso
l’interno.
Sughero, fellogeno e felloderma vanno
a costituire il PERIDERMA
Legno secondario
Il legno secondario prodotto dall’attività
del cambio cribro vascolare svolge
principalmente 3 funzioni:
1. Lo xilema prossimo al cambio,
recentemente differenziato, assicura
il TRASPORTO dell’acqua e sali in essa
disciolti.
2. Allontanandosi dal cambio, andando
verso il centro, si inizia a perdere
gradualmente la funzione di trasporto
e inizia quella di SOSTEGNO (tille,
3.
protuberanze
delle
cell.
parenchimatiche, occludono i vasi ed
entra
aria
facendo
perdere
funzionalità).
RISERVA: le cellule parenchimatiche
durante
il
riposo
vegetativo,
immagazzinano le sostanze elaborate
dall’azione fotosintetica. Il legno +
interno, + vecchio, non svolge questa
funzione, perché qui le cellule sono
morte.
La sezione di alcuni tronchi (Prunus, mogano, teak, ebano…)
presentano 2 zone facilmente distinguibili:
• Alburno: esterno e prossimo al cambio cribro vascolare, è la
zona+ chiara: è lo xilema funzionante nel trasporto dell’acqua e
svolge funzione di riserva per le cellule parenchimatiche vive.
• Duramen: interna e lontana dal cambio cribro vascolare e
costituita dal legno più scuro, più vecchio. Lo xilema non è più
funzionante, le pareti cellulari sono impregnate di tannini, oli
essenziali, resine che sono antidegradative e antifermentative,
che non fanno marcire il legno e danno ad esso un colore
particolare, ne incrementano la durezza, il peso l’impermeabilità.
È quello commercialmente + pregiato.
In alcuni alberi, mancano queste impregnazioni del duramen (salice)
ed il tronco può per l’azione di funghi, divenire cavo. Nell’ Acer
pseudoplatanus, Acer platanoides ad esempio è presente solo
l’alburno.
Libro secondario
Il floema funzionante è solo quello dell’ultimo anno, più
prossimo al cambio.
Esternamente a questo troviamo del tessuto floematico
che mantiene solo la funzione di riserva.
In ogni caso, lo spessore totale del floema è sempre
ridotto in quanto, con il procedere dell’accrescimento
diametrico della pianta, esso viene schiacciato verso i
margini del fusto e di esso rimangono solo le fibre.
Anche qui il lume degli elementi cribrosi viene occluso da
un callo in modo da divenire non funzionale.
A seguito dell’attività del fellogeno, inoltre, il vecchio
floema viene eliminato con la caduta del ritidoma.
Nelle piante erbacee, l’asse caulinare non
possiede cellule con pareti lignificate e
prende il nome di fusto. Negli arbusti e
negli alberi al contrario l’asse lignifica,
diventa rigido e prende il nome di
tronco.
La porzione dell’asse compresa fra il
colletto e le foglie cotiledonari viene
detta asse ipocotile. La prima parte del
fusto che si forma dopo la germinazione
.
Fra i cotiledoni e la prima foglia è situato
l’asse epicotile.
Il punto di inserzione della foglia sull’asse
è spesso ingrossato e va a formare un
nodo. La porzione di fusto fra due nodi
consecutivi si dice internodo.
La
crescita
intercalare
dovuta
ai
meristemi permette l’allungamento del
fusto e l’allontanamento dei nodi. Il
fusto dove gli internodi hanno raggiunto
la loro massima lunghezza viene detto
macroblasto.
Nelle piante primitive la ramificazione
era biforcata (dicotomica). La
ritroviamo ancora nelle briofite e
nelle pteridofite attuali(Psilotum).
Ha inizio a livello di una cellula apicale
che si divide longitudinalmente in
determinati momenti
dell’accrescimento del fusto. I due
rami della forcella si sviluppano a
partire da due cellule figlie.
I fusti delle spermatofite, danno
origine a ramificazioni che nascono
dalle gemme laterali (o ascellari).
Una gemma corrisponde ad un cono
vegetativo racchiuso da foglie
modificate con funzione protettiva.
Le gemme laterali sono situate sul
fusto all’ascella delle foglie
Le foglie possono presentare
all’ascella una o + gemme a
seconda della specie.
Le ramificazioni del fusto si
sviluppano da gemme ascellari.
Se un ramo laterale non si allunga
viene detto brachiblasto.
In parecchie specie arboree, ad es.
negli alberi da frutta si
osservano brachiblasti che
portano fiori e successivamente
frutti. Questi rami sono detti
fruttiferi.
Le foglie di larici e pini sono alla
stessa stregua inserite su
brachiblasti.
Il fusto di numerose piante legnose
si allunga con macroblasti
durante il primo anno mentre
l’anno seguente si sviluppano
brachiblasti laterali
Le modalità con le quali il fusto può ramificarsi possono essere raggruppate
essenzialmente in due tipi principali:
Ramificazione monopodiale
Ramificazione simpodiale
La forma del cono è determinata dalla posizione delle gemme laterali che si
svilupperanno.
Si distinguono un accrescimento: Acrotono e Basitono
Le regole che si applicano alla ramificazione del fusto valgono anche per le
infiorescenze
Le modalità con le quali
il fusto può ramificarsi
possono
essere
raggruppate
essenzialmente in due
tipi principali:
Ramificazione
monopodiale
Ramificazione
simpodiale
La forma del cono è
determinata
dalla
posizione delle gemme
laterali
che
si
svilupperanno.
Si
distinguono
un
accrescimento:
Acrotono
Basitono
Le regole che si applicano
alla
ramificazione
del
fusto valgono anche per
le infiorescenze
Gli anelli di crescita
Molto evidenti, nelle sezioni trasversali di fusti di piante che vivono nelle zone temperate, sono
un gran numero di anelli concentrici chiamati anelli di crescita. Osservando la struttura di
questi anelli al microscopio ottico, si può notare che essi si originano in quanto le cellule
conduttrici dello xilema, non hanno tutte lo stesso diametro.
Ad elementi conduttori con lume molto grande, fanno seguito, procedendo dal centro verso il
cambio c.v. elementi conduttori dal diametro progressivamente minore, evidenziabili come
zone più scure perché con minore contenuto di aria, finché ad elementi molto piccoli,
seguono bruscaente elementi con diametro molto grande.
Ciò si spiega in quanto il cambio di piante legnose di zone temperate, alla ripresa in primavera
dell’attività vegetativa, sotto lo stimolo di ormoni prodotti dalle gemme, produce elementi
con pareti sottili, facilmente estensibili e dal diametro grande, per garantire l’appropriato
apporto di acqua alle gemme che si devono aprire.
Con il sopraggiungere dell’estate, invece, il cambio inizia a produrre elementi elementi sempre
più piccoli e con pareti fortemente ispessite e lignificate, finché al cessare dell’attività
meristematica del cambio (in genere ad Agosto), si interrompe la produzione di nuovo
xilema. Nella primavera successiva, il cambio riprende a produrre elementi grandi che, a
stretto contatto con quelli piccoli e con pareti spesse dell’anno precedente, creano un
confine netto tra il legno primaverile e quello estivo. Ogni anno un nuovo anello si aggiunge
ai precedenti.
La formazione di un legno estivo diverso da quello primaverile, non è tanto dovuta alla
mancanza d’acqua in estate quanto ad una variazione, sia qualitativa che quantitativa, nella
produzione di ormoni dalla parte delle gemme, che in questo periodo cessano di svilupparsi.
Se si eliminano le gemme apicali di una pianta, l’attività del cambio cessa, per riprendere se al
suo apice si usano ormoni come le giberelline o le auxine. Esiste quindi uno stretto rapporto tra
la formazione di foglie e l’attività del cambio.
Ovviamente il fenomeno degli anelli di crescita è presente solo in quelle piante che vivono in
zone dove il cambio è attivo periodicamente e dunque dove esiste l’alternanza delle stagioni.
Nelle zone equatoriali, dove la temperatura è praticamente costante durante tutto l’anno, il
cambio è sempre attivo e non si hanno quindi anelli di crescita. Il legno privo di anelli è molto
pregiato ne è un esempio l’ebano. Se in una stagione vegetativa (corrispondente ad un anno
nelle regioni temperate) si forma un solo anello di crescita, questo viene detto anche cerchia
annuale.
Epidermide
Sezione trasversale di foglia di Nerium Oleander
È in genere monostratificata
(eccezione xerofite-oleandro)
Tessuto complesso che riveste solo
la porzione epigea della pianta.
Le cellule che costituiscono
l’epidermide sono varie per
forma e funzione:
1. cellule epidermiche
2. cellule di guardia
3. peli o tricomi